Phylogenetic status, characteristics and geographical distribution
Pre-Pleistocene Fossil Primates
Dryopithecus:
- Early fossil finds of Dryopithecus (first definite discovery in 1856) were mainly teeth. Today we have a cranium and many log bones as well. This has not only increased our subject knowledge about Dryopithecus but also has helped to scientifically check earlier conclusions. Dryopithecus is considered to be a common ancestor of both man and modern apes.
- On the basis of earlier evidences it was believed that Dryopithecus survived from middle to the end of the Miocene period.
- Y-5 cusp pattern on his 3rd molar indicates that he was ancestor of both man and modern apes. His other body features are also similar to modern apes.
- His three species i.e. africanus, major and sivalensis must have given rise to three modern great apes of today that is Chimpanzee, Gorilla and Orang.
- Recent evidences however indicates that D. sivalensis showed absolute similarities with Ramapithecus and therefore the two has been clubbed on to a new genus Sivapithecus. His first lower premolar is sectorial that separates him from later hominids.
- In 1948 Mary Leaky found a preserved cranium which was named Proconsul but now it is considered as a member of Dryopithecus africanus.
- In comparison to present day apes, his jaws, teeth and muscles were similar and many features of his skull resemble monkeys.
- Recently walker studied his skull and estimated the weight of D. africanus to be 26 pounds whereas other anthropologists consider it to be slightly heavier i.e. approx. 33 pounds.
Dryopithecus: Important Facts
- Time span : 23m to 8m years ago.
- Arboreal habitat- both rain forests and more open areas.
- Size variable, from gibbon to gorilla.
- Large teeth, interlocking canines, Diastema.
- Y-5 Cusp Pattern.
- Limbs- mixed characteristics of both apes and monkeys.
- Indication of ape simian shelf.
- Not modified for knuckle walking.
- Evolutionary status- possible ancestor of chimpanzee, gorilla and man.
- Geographical distribution - Africa, Asia and some parts of Europe.
Walker and Tifford estimated its cranial capacity to be about 167 c.c. According to this, in proportion to body size the brain size of africanus was more than that of monkeys.
On the basis of presence of frontal sinus, it can be concluded that he was related to man and African apes.
Post-cranial fossils of Dryopithecus found in the last decade show that the many characteristics of his limbs resemble that of modern apes and monkeys. On the basis of the study of these parts it is concluded that africanus was slow walking animal and lived on trees and his limbs were not modified for Brachiation and climbing.
Ramapithecus
Ramapithecus is the most important hominid from Miocene period. In recent years Ramapithecus has been accepted by many scholars as the first true hominid. There are at least two dozen fossils specimens that have been identified as belong to Ramapithecus. Most of these specimens consist of teeth and jaws and they principally come from two areas - the Siwalik Hills in India and Fort Ternan in Kenya.
Discovery and Distribution of Ramapithecus:
The first discovery of Ramapithecus fossils was made by G.E. Lewis in 1932 in the Siwalik hills regions of India. He assigned one of the fossils, an upper jaw, to a new genus and species he named Ramapithecus brevirostris.
The generic name simply means Rama's ape' Rama being the mythical prince who is the hero of Indian epic poem. The species name that Lewis chose was more meaningful for it is the Latin word for 'short snouted'.
Next Ramapithecus fossil find was made by L.S.B. Leakey near Fort Ternan in south western Kenya in 1961. The specimen included parts of both sides of an upper jaw. Leakey gave it the name Kenyapithecus wickeri, which is synonymous with Ramapithecus brevirostris, after his friend Fred Wicker, on whose farm the fossil was found. which is synonymous with Ramapithecus brevirostris.
The next Ramapithecus specimen was excavated by Von Freyburg, a German geologist, in Greece during World War II.
The specimen was assigned to another new genus and species: Graecopithecus freyburgi. Freyburg's find was the complete tooth bearing part of lower jaw and at the time of its discovery it contained all the teeth.
Next to the growing inventory of Ramapithecus fossil was a lower jaw unearthed from a Miocene deposit near Candir, some 40 miles north east of Ankara in Turkey in 1973.
The specimen was named as Sivapithecus alpani. The species name of the Candir jaw honors the director of the Turkish Geological Survey.
Anatomical Characteristics of Ramapithecus
- Incisors and canine are inserted vertically and not in slight procumbent position as in apes.
- Little or no canine diastema.
- The canines of the Ramapithecus are not projected and they possess narrow faces.
- The dental arcade is rounded.
- The palate of the Ramapithecus is arched as in man.
- Flattened and thick enameled premolars and molars that appear to be adapted for heavy chewing and processing of hard food stuffs.
- Ramapithecus has a canine fossa ( Kenyapithecus).
- The molars possess the Dryopithecus Y-5 cusps pattern.
- Slightly divergent tooth rows. The tooth rows have been identified as parabolic by some and V-shape by some others.
- Reduction of size of third molar as compared to first and second molar.
- The ratio between the sizes of front tooth (incisors and canine) and those of cheek teeth (premolars and molars) is roughly the same which indicates the human position.
- Shelf-like ridges are present inside the lower jaw of Ramapithecus.
- Large inferior torus on mandible.
- Short maxilla that would indicate a placement of the chewing muscles that increase the chewing pressure brought to bear on the food being eaten.
PHYLOGENETIC POSITION OF RAMAPITHECUS
To understand phylogenetic position of Ramapithecus we have to take back our observations back to Dryopithecus first.
The Dryopithecinae primates made their appearance in Europe, Asia and Africa during Miocene and Pliocene epochs. Their size ranges from gibbon like body form to the body structure of modern gorilla. Most of the remains which belong to Dryopithecinae are jaws and teeth.
Therefore, the characters distinguishing Dryopithecinae from Hominidae are restricted to dentition. Gregory and Hellman, after conducting their dental characters, came to the conclusion that Dryopithecinae were the common ancestor of the anthropoid apes and man.
In the year 1856, Lartet discovered from Miocene deposits, in south France, a lower jaw bone which was assigned to the genus Dryopithecus. The place of Dryopithecus in the evolutionary stem has been found out by studying the peculiar dentition - "the Dryopithecus pattern" which is characterized by five cusped lower molars. After careful study of the different species of Dryopithecus, it has been decided by many scientists that Dryopithecus fontani, Dryopithecus rheuanus and Dryopithecus darwini, were probably the ancestors of gorilla, chimpanzee and humanoid forms respectively.
Fossils found in Europe and Asia since 1970 suggests that between 10 and 15 million years ago Dryopithecus gave rise to at least three other genera. Two of them Sivapithecus and Gigantopithecus were primates with a face as large as that of a modern chimpanzee or gorilla.
The third genus, Ramapithecus had a small face. Of the three genera, Ramapithecus clearly shows the greatest similarity to later hominids.
Ramapithecus has been the center of a great deal of debate concerning its possible hominid status. Pilbeam has proposed alternatively that a number of the middle and late Miocene genera to be classified together in Ramapithecines, in an attempt to both draw attention to morphological feature shared by the group which differentiate it from others and to focus discussion on adaptation and biology rather than phylogeny.
The most widely distributed Ramapithecids genera are Ramapithecus and Sivapithecus. The taxonomy of this group is in a rather confused state, which newer materials from Pakistan will hopefully help clarify. Isolated teeth of Ramapithecus and Sivapithecus are very difficult to distinguish except on the basis of size; Ramapithecus teeth are smaller.
What seems more probable is that both Ramapithecus and Sivapithecus are quite dimorphic dentally and that the size ranges of the two forms overlap, perhaps substantially Ramapithecus may show less canine dimorphism than Sivapithecus though more than the Pliocene hominid Australopithecus afarensis.
A handful of Ramapithecids postcranial remains have been recovered during recent work in Pakistan, attributable to Ramapithecus, Sivapithecus and to a third form Gigantopithecus bilaspurensis. Though these remains are unfortunately fragmentary, they suggest that all the Ramapithecids were smaller than previously expected: Ramapithecus ~20kg, Sivapithecus - 40kg and Gigantopithecus - 70kg.
Therefore, the fossil finds of Ramapithecus are regarded as the most important addition to the knowledge of relating to human evolution. Credit goes to G.E. Lewis to discover in the year 1934, the fossilized remain of Ramapithecus in the Siwalik hills of India. Dr. Simons has attributed Ramapithecus a very significant position in the line of human evolution.
Ramapithecus raised many important points which are highly effective in search of human ancestral pattern. On examining the nature and extent of teeth, some scholars described Ramapithecus as a weapon wielding terrestrial biped. Ramapithecus, according to the competent anthropologists, represents the oldest known ancestors of the human line.
The scientists like Simon, Pilbeam and Tattersall are the proponent of Ramapithecus as a human ancestor. The materials so far excavated in relation to Ramapithecus suggest a line between Dryopithecus group belonging to early Miocene and later real hominids. In a review based study made by Conroy and Pilbeam a plausible interpretation of the Ramapithecus has been given as the late Cenozoic ancestor of Australopithecus.
In consequence of recent findings and interpretations Ramapithecus has been widely considered as a candidate for the first hominid. It splits up from the ape line 14 million years ago and marked the remarkable beginning of hominid line.
The main reason for giving Ramapithecus a true hominid status is the similarity of its teeth with that of the later hominids. In discussing the status of Ramapithecus, Swartz and Jordan have remarked that when a creature is called hominid, it doesn't mean that it is a modern man, but this term is used for clearly human like forms. Ramapithecus was such a creature as understood by many authorities.
Controversy Regarding the Taxonomy of Ramapithecus
The current view of the Ramapithecus depends upon little more than two dozen fragments, mainly of teeth and parts of jaws that have been discovered since the first find reported on by G. Edward Lewis in 1934.
The initial discovery prompted Lewis to recognize a new form that he called Ramapithecus. This was followed in later years by a handful of fossils that were each recognized as new forms and they were given a series of separate names (Kenyapithecus, Graecopithecus, Rudapithecus, Sivapithecus) based upon the geographical localities at which they were found.
But in 1965 Simons and Pilbeam reviewed the entire series and held the view that all these forms really comprised two species groups. One of these, Sivapithecus, was basically ape like and it was therefore put forward as an ape ancestor; the other, Rudapithecus, seemed to possess a number of hominid- like features was therefore entered as an early hominid ancestor. This view was still extent in 1977 but a series of more recent studies has cast doubt upon it.
Thus Andrews and Cronin (1982) and Lipson and Pilbeam (1982) have all suggested that the non-Chinese Ramapithecus are really only a single species or species group, that the two forms ( Sivapithecus and Ramapithecus) are really only the males and females of sexually dimorphic species group.
One of the reasons for putting forward this new idea is an attempt to make these data conform to those suggested by the concept of molecular clock.
The molecular clock, assessing the time from common ancestry of two species using the notion that molecular evolution has taken place in a linear manner, suggests that human and African apes had a common ancestor at five million years ago or even closer to the present time.
If these were true, it would be logically impossible for there to have existed prior ancestors of humans (Ramapithecines date from 8 to 14 million years ago) that were more like humans than apes. The new views of the fossils have therefore concentrated on the ape like features of Ramapithecines and of these, big sexual dimorphism is one of the most powerful, being found in every great ape known, but not markedly present in any species of the genus Homo so far identified. But the later evidences regarding Ramapithecus strongly suggests that two species are present there in Yunnan. One of these, the larger creature, (Sivapithecus), with larger dental sexual dimorphism, larger canine dimorphism, larger canine heights and areas, more herbivorous dentition, considerably smaller number of males than females has attributes that are matched by many of the apes.
In contrast, the smaller creature, (Ramapithecus) possess smaller dental sexual dimorphism, smaller canine dimorphism, smaller canine heights and areas, more omnivorous dentition and equal numbers of males and females, and thus has attributed that would not deny it a place in a radiation of prehumen form.
Gigantopithecus
History of Gigantopithecus is as interesting as its fossils and its evolutionary status. For centuries China's pharmacies used bones of giant animals to make medicines.
In 1935 Koenigswald, in one of such medical stores found such remains and named them Gigantopithecus. Paleontologists/evolutionary biologists differ regarding its evolutionary status. Some consider it as giant ape, others as snowman and ancestor of Yeti of Himalayas. At present many fossils of Gigantopithecus - many mandibles (broken to almost complete) and more than a thousand teeth have been found from China, India and Vietnam.
On the basis of remains according to Simons and others, Gigantopithecus was about 9 feet tall and weighed around 600 pounds. Though this conclusion has possibly some boosting but still size of this giant like genus can be approximated. Possibly Gigantopithecus was 6-9 feet tall and weighed 450-600 pounds. His Indian species bilaspurensis (discovered by late prof. S.R.K. Chopra of Punjab University) has been dated from 9 million to 5 million years ago whereas Chinese and Vietnamese species blacki existed in middle Pleistocene period.
Gigantopithecus had large sized teeth and solid/hard enamel. Jolly, by his seed eater hypothesis has tried to prove that structure of teeth is based on their dietary habits & Gigantopithecus food should be dominated by hard nuts and other solid fruits.
Many anthropologists don't agree with Jolly's view, but one thing is sure, that this middle Pleistocene quadruped cannot be the ancestor of the man. Probably around 5 lakh years ago it became extinct.
Gigantopithecus : Main Features.
- Time span : 9 million to about 0.5 million years ago
- Large sized
- Large sized teeth, interlocking canines, diastema
- Dental arcade U shaped
- Species - blacki, bilaspurensis
- Geographical distribution - India, Vietnam and China
- Evolutionary status - not an ancestor of man, became extinct.
Evolutionary Status of Miocene Hominoids
Evolutionary status of Miocene Hominoids and Hominoids before Miocene is not clear even today.
Dryopithecus is the first evidence of ancestry of modern apes. This ape resembled modern apes in dentition but regarding rest of the physical features and locomotion it can be compared more with monkeys.
The first clear evidence of ancestry og modern apes is clear through Aegyptopithecus but situation after this is not clear. There are many variations. Types on fossils of Dryopithecus. Because of this it is difficult to know regarding which of the Dryopithecinae with certain definite characters evolved towards direction of human evolution.
Status of Sivapithecus as ancestor of orang is clear. Gigantopithecus was too specialized aan ape and has no place in the ancestry of man.
Status of Gigantopithecus vis a vis other hominoids of the period is not clear. On the basis of biochemical studies of blood proteins of Primates in recent 2-3 decades, it has been concluded that evolutionary line of gibbon separated from human line about 30 million ago, whereas till about 8-6 million years ago the man, chimpanzee and gorilla had a common ancestry.
On the basis of DNA studies etc. in last decades it can be said that direction of evolution of man and apes separated between 8 million and 6 million years ago. Unfortunately, hominid fossils of late Miocene period have not been found yet. So it is difficult to conclude who was the ancestor of later definite hominids.
Australopithecine
Most of all the Hominid fossils of Africa comparing with the African apes have large brain, relatively reduced chewing or molar teeth and limb bones that are, except in points of details, like those of modern Homo sapiens. However, there are at least four and perhaps five species of hominid that lack expanded brain, the reduced molar teeth and the skeletal features of Homo. They antedate or overlap with earliest representatives of Homo and are known collectively as the Australopithecines. Hence, we should understand importance of this hominid fossil (Australopithecines) in tracing the evolutionary line of present day human being.
Raymond Dart first discovered in 1924 at Taung in South Africa. The finding includes part of mandible, facial skeleton and endocranial parts of a child. Dart named the skull as Australopithecus africanus.
The meaning of austral is south and pithecus means ape. At first discoverer of the various
Australopithecine fossils named four to five genera, but now the consensus of scientific opinion recognizes one genus Australopithecus and two species, a slenderly built and smaller form, Australopithecus africanus (Gracile form) and larger roughly built Australopithecus robustus originally called Paranthropus.
The following are the fossils along with the sites found in South Africa and East Africa:
|
Place
|
Fossils |
Discoverer |
|
Taung |
Australopithecus africanus |
Raymond Dart, 1925 |
|
Kromdraai |
Paranthropus robustus |
Robert Broom, 1938 |
|
Makapansgat |
Australopithecus promethecus |
Robert Broom, 1948 |
|
Stenfontein |
Plesianthropus transvalensis |
Robert Broom, 1936 |
|
Swartkrans |
Paranthropus crassidens |
Robert Broom, 1947 |
Fossils from East Africa:
|
Place
|
Fossils |
Discoverer |
|
Hadar |
Australopithecus afarensis (Lucy) |
C. Johanson, 1974. |
|
Omo |
Australopithecus africanus,
Australopithecus boisei, Homo habilis and Homo erectus |
|
In East Africa, Louis Leakey made the first discovery of a hominid fossil from Bed I of Olduvai Gorge in 1959. The first discovery named Zinjanthropus boisei, now known as Australopithecus boisei is considered to be a super robust Australopithecus. Another important discovery was announced in 1964 from Olduvai Gorge and named Homo habilis.
Other important sites in East Africa are the Omo river valley of Southern Ethiopia where both robust and gracile forms have been found. Similarly both robust and gracile forms were discovered from Hadar in Eastern Ethiopia and Turkana region of Northern Kenya. Turkana region have been studied by Richard Leakey since 1969 and over 120 hominid fossils have been discovered so far.
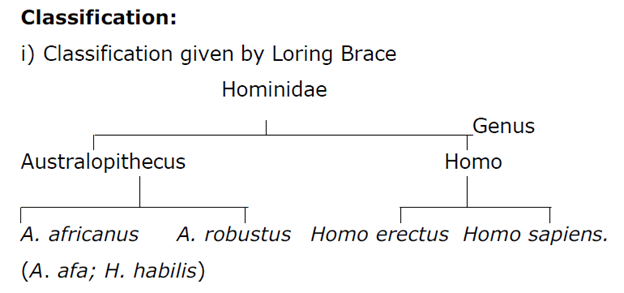
According to Loring Brace Homo habilis is the member of Australopithecus africanus but for John Robinson it is not.
T. Robinson's Dietary hypothesis:
Going by the dental fossil evidence found in East and South Africa, there are two adaptive groups which can be extended to families. These two adaptive groups are gracile and robust. They are quite distinct in morphological, ecological relationship and behavior. Gracile group is Australopithecus whereas Robust group is Paranthropus.
He sees a dichotomy in cranio - dental form separating these two Australopithecine groups. Robust group has the large post canine teeth crowns and its thick canine has larger occlusal surface, well developed roots and relatively flat occlusal surface (greater enamel has been damaged). Crownding and size reduction of the front teeth, point to a primary dietary function of crushing and grinding.
The massiveness of the entire masticatory apparatus including the musculature and the wear of the teeth indicate a diet of rough material which needed much chewing, hence robust was herbivorous or vegetarian. Gracile forms show none of these features, rather their post canine teeth are smaller and anterior teeth would be substantial until tool making had reached a fairly advanced level.
Age and Antiquity of place:
The Taung cave once considered to be very ancient actually seems to be the most recent one, less than 0.9 million years ago. The specimen may represent one of the last surviving members of the genus Australopithecus. The absence of suitable materials precludes the use of Radioactive dating in South Africa.
In Olduvai Gorge pre historic volcanic activities provided abandoned material for the application of Radioactive dating techniques. The Australopithecus remains are dated by potassium argon method and placed between 1.9 and 1.1 million years. Sites of the Omo valley has been dated between 2.75 and 2.5 million years, the Hadar material is dated between 3 to 3.5 million years and the Lake Turkana is around 3 to 1 million years. However gracile Australopithecus from other sites of East Africa such as Middle Awash and Lothagam extend the age range of this group of fossils to 4 million years and beyond perhaps to 5 million years.
Characteristics of Australopithecus africanus:
Skull:
Taken as a whole, the skull is a combination of small brain case and large jaws. The brain case lacks high vertical forehead of Homo sapiens and the high roundedness of the skull vault. This gives a simian appearance. But in bone feature it contrasts with pongidae and matches with
hominidae.
- The supraorbital height index shows that the relative high exceeds range of variation in anthropoid ape and actually comes within the range of hominid skull.
- The occipital torus and the inion occupy a low level as in hominid skulls. In adult apes this occipital torus forms a crest high up the occipital aspects of the skull, thus extending considerably the nuchal area for attachment of neck muscles. In Australopithecus the nuchal area is restricted as in homo.
- In Australopithecus the occipital condyles are forward in position relative to the total length of the skull and the auditory aperture. The occipital condyles of pongidae are behind the midpoint of the cranial length and also behind the auditory apertures.
- In all the Australopithecus skulls in which mastoid region are sufficiently well preserved, there is a well marked pyramidal process typical of hominid form.
- The brow ridges are poorly developed compared to that of the apes.
- The facial skeleton is however, large in relation to the brain case.
- The cranial bones of Australopithecus robustus are thicker than the Australopithecus africanus and the sagittal crest is developed. The zygomatic arches are expanded and flared.
- The jaws are also relatively large but no simian shelf.
Cranial capacity:
In cranial capacity, the range of variation is quite considerable. The skull of Australopithecus afarensis and Australopithecus africanus ranges from 400 to 500 cc and Australopithecus robustus ranges from 410 to 530 cc.
Thus the general endocranial size of Australopithecus does not differ markedly from those of gorilla and chimpanzee. However in relation to the body size Australopithecus shows larger brain capacity proportion than the apes.
Dentitions:
Australopithecus dentition is essentially of hominid type. In all the adult specimens the dental arcades is evenly curved as in Homo sapiens with no diastimic intervals. The canine is reduced in size and spatulate in forms. Canines and incisors are almost in the same level.
The anterior upper premolars have two roots, the anterior lower premolars are non-sectorial and have one root (sectorial and two roots in apes). There is good evidence for the study of the immature specimens that the order of eruption of permanent teeth agrees with the Hominidae. In Homo sapiens the canine erupts before the second molar. This is reverse in the apes.
Pelvis:
The total morphological pattern of the pelvis of Australopithecus is hominid.
- The anterior inferior iliac spine is strongly developed.
- The ilium is much relatively broader than in the apes. The broad ilium lengthens the attachment of the gluteus muscles that makes important in maintaining balance of the trunk on the legs. The gluteus maximus becomes a fourfold extension muscle which is needed for erect bipedal locomotion. Whereas in monkeys and apes the gluteus maximus is an abductor muscle.
- The posterior extremity of the iliac crest is extended backward and downward in the sacral area.
- The angle of the sciatic notch (depression) is more acute in the pelvis of man and Australopithecus than that of the great apes. This is resulted in part due to development of prominent ischial spine.
- Ischial tuberosity is relatively high in man and Australopithecus and closer to acetabulum than that of the apes.
- Sacrum is shifted upward and closer to the acetabulum forming more or less a basin or funnel shaped pelvis which indicates the character of erect posture.
Limb bones:
In addition to the pelvis, the Australopithecus finds include many fossilized bones of leg, hand and feet. One group from Olduvai Gorge includes twelve of the major bones of a single foot and so perfectly preserved that the details of the foot are readily reconstructable.
The foot of Australopithecus is much smaller than a human foot. But the metatarsals are hominid and the big toe points forward and not splayed out. The carpe bones and phalanges found in Olduvai Gorge also show hominid characters.
Two specimens of the lower part of femur found at Stenfontein have been described. They show a combination of features such as the obliquity of the shaft, the alignment of the condyles, the forward prolongation of the intercondyler notches which confirm to the hominid femur.
Upper extremity of one specimen, however shows certain Pongidae features, such as relatively small size of the articular head. A tibia and fibula found in Olduvai have relatively straight shafts conforming to hominid character.
Difference between Australopithecus africanus and Australopithecus robustus:
|
Character
|
Australopithecus africanus |
Australopithecus robustus |
|
Height |
1.1 - 1.4 m. |
1.2 - 1.4 m. |
|
Weight |
30 - 60 kg. |
40 - 80 m. |
|
Physique |
Light build, probably relatively long arms, more human
features; Probably less sexual dimorphism. |
Very heavy build, relatively Long arms; marked sexual
dimorphism. |
|
Brain size |
400 - 500 cc |
410 - 530 cc |
|
Skull form |
Low, flat forehead, brow ridges less prominent. |
Prominent crest on top and back of skull, long broad flattish
face strong facial buttressing. |
|
Jaw teeth |
Small incisor like canine, no gap between upper incisors and
canines, larger molars. |
Very thick jaws, small incisors and canine, incisors and
canine, very large molars. |
Phylogenetic Position:
Currently there are three main phylogenetic trees each with its own cadre of proponents. The problem of whether Australopithecus africanus represent early grade in homo lineage or a small brain hominid arising as a separate product of Ramapithecus radiation is yet to resolve.
At the same time we also do not know how the two dimorphic forms i.e. gracile and robust Australopithecines are to be classed phylogenetically. Whether they reflect polytypic nature of single taxon or systematic enough to conceive as two taxa is still a question.
There is no generally accepted phylogenetic tree for human evolution. There are several proposal branching perhaps the major distinction between them is whether homo is perceived as late arrival or as early arrival.
In some phylogenetic, robust is consider to be on the same evolutionary line with Australopithecines and some consider robustus to be separate from the earlier gracile species in which Paranthropus robustus/promethecus is equivalent to Australopithecus robustus and Homo africanus equivalent to Australopithecus africanus.
This Pliocene and Pleistocene fossils have shown both hominid and ape like features. However the hominid features are overwhelming. The presence of ape like features can be accounted for by way of common inheritance from a hominoid or Pongidae ancestor.
The hominid characters however can be accounted for by way of independent acquisition demonstrating and highlighting the fact that these fossils were on a direct line of human evolution and not Pongidae line.
There are many schools of thought prevailing with reference to the course of evolution these australopithecine have taken. The most important of these are discussed below.
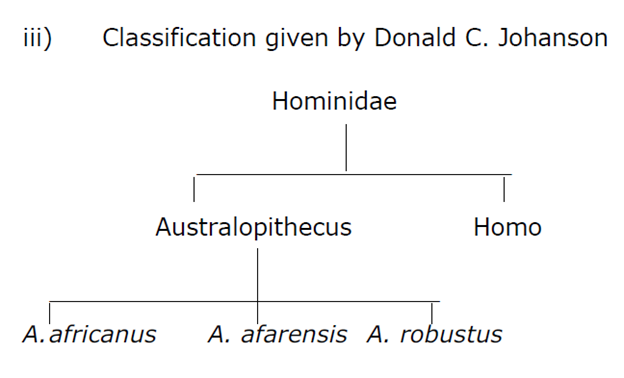
According to Donald C Johnson and Timothy white the east African fossils Australopithecus afarensis split into two branches and australopithecine line represented by A. africanus P. robustus, P. boisei and a hominid line represented by homo habilis, homo erectus and homo sapiens.
This split was supposed to have happened 3 million years ago. The australopithecine line progressively became robust. This pattern generally called two branched theory had its variants also. For some it is A. africanus which is the common link between the australopithecine line and homoline. For still others these two branch represent parallel evolution.
According to this two branch theory A. afarensis gave rise to A. africanus 3 million years ago of the same height living up to two million years ago.
Next arrived A. robustus which showed marked increased in the robustness of the body face , jaws and teeth and it had lined up to 2.3 to 1.8 million years ago. Finally the last and the most robust form A. boisei lived in east Africa from roughly 1.8 to 1 million years ago.
The second branch of this model the homo line also shows a shortening of the face but there is marked decrease in the size of both cheek teeth and the front teeth. There is a massive increase in the size of brain also. This line begins with ta transition form A. afarensis to H. habilis the first hominid who made and used tools and lived in Africa from 2-1.5 million years ago. They had human like teeth and larger brain than australopithecine (750 cc). however we can conclude that hominid evolution may not have been so simple, isolated and clear cut in tis operations.
Instead there is a possibility that three or more hominid lineages may have been evolving and interacting with each other.
This two branch theory was widely accepted till the discovery of the new type of hominid skull - Australopithecus aethiopithecus in northern Kenya inn 1985 by Alan Walker. This skull is considered to be the most robust form ever discovered. It had massive teeth and ape like brain.
The dating of the this specimen indicates that the family of a. boisei didn't evolve in the last leg of the australopithecine evolution as indicated by earlier theory but it originated directly from a. afarensis. Thus the revised theory holds a three line evolutionary sequence. One to boisei line second to Homo and third to africanus, robustus line having A. afarensis as the common ancestor.
The discovery of the youngest australopithecine - A. ramidus has added a new dimension to the three branch theory. It holds that A. afarensis is the common ancestor of Homo, P. boisei and P. robustus but it itself evolved from A. ramidus. With overspecialization of diet, competition for food with H. habilis and the latter's predation along with H.erectus led to the extinction of australopithecine group.
Why are the genera Australopithecus divided into two species?
At first, several discoverers of the various Australopithecus fossils named four to five different genera, but now the consensus of scientific opinion recognizes one genus Australopithecus and two species, one gracile form represented by Australopithecus africanus and other robust form represented by Australopithecus robustus, originally called Paranthropus. This division is made on the basis of their morphological feature.
The gracile form are slenderly built and smaller; gracile form have probably relatively long arm, probably less sexual dimorphism, high forehead, shorter face, brow ridges less prominent. The incisors are also small with no gap between upper incisor and canines and have big molars; cranial capacity ranges from 410-530 cc.
The robust form are large and roughly built. They have relatively long arm with moderate sexual dimorphisms, crest on top of skull, flattish face. The robustus have thick jaws, small incisors and canine, very large molars; the premolar is also large like molars. The cranial capacity ranges from 410- 530 cc.
What is the position of Australopithecus in the line of human evolution?
Different scholars have given different opinion about the position of Australopithecus in the line of human evolution. According to Johanson and White Australopithecus afarensis is regarded as the common ancestor of Australopithecine and Homo.
Australopithecus afarensis rise to Australopithecus africanus and to Homo in one hand and directly to Australopithecus robustus in the other hand. Here Australopithecus africanus act as a transition form from Australopithecine to Homo.
According to L.S.B. Leakey, the common ancestor of both homo lineage and Australopithecine divided quite early. He considers Australopithecus afarensis as not the common ancestor. The homo lineage starts separately after 3 million years and Australopithecus a little bit early but also only 3 million years.
What is T. Robinson Dietary hypothesis?
Going by the dental fossil evidence found in East and South Africa, there are two adaptive groups which can be extended to families. These two adaptive groups are gracile and robust. They are quite distinct in morphological, ecological relationship and behavior.
T. Robinson sees a dichotonomy in cranio-dental form separating these two Australopithecines groups. Robust group has the large post canine teeth crowns, thick enamel, larger occlusal surface, well developed root and relatively flat occlusal surface (greater enamel has been damage).
Crowning and size reduction of the front teeth points to a primary dietary function of crushing and grinding. The massiveness of the entire mastigatory apparatus including the musculature and the wear of the teeth indicate use of a diet of rough material which needed much chewing.
Hence robust was herbivorous or vegetarian. Gracile form shows none of these features rather their post canine teeth are smaller and anterior teeth larger. The needs for these in meat eating would be substantial until tool making had reached a fairly advanced level.
Homo Habilis
The first H. habilis, described in 1964, came from Oluvial Gorge in Tanzania. The original material was fragmentary, sufficiently so to provoke some controversy over whether a species distinct from south African A. africanus was truly present; H. habilis was said to have had a larger brain and smaller cheek teeth. Since then, more material has been found at Olduvai, Omo and east Turkana where several relatively complete crania were found including ER 1470 from east Turkana. The age of ER 1470 was originally thought to be in excess of 2.6 million but it is now dated around 2 million years.
Homo habilis consist of several reasonable crania, some jaws and teeth, and a few unassociated post-cranial. There are disagreements over the number of species being sampled. If it so only one there is a considerable range in rain volume, A small number of larger crania cluster around 500 cc. These group differ in tooth size, the former having larger teeth including canines and facial morphology differs markedly between the best preserved representative of each group, ER1470 and ER1813 both from east Turkana. If these are males and females the degree of brain, tooth and face size dimorphism is considerably greater than in any living species.
We assume here that two species are probably being sampled, Homo habilis and smaller brained species superficially resembling A. africanus. Homo habilis specimens are known from a little before to little after 2 million year ago from east Turkana, Olduvai, probably Omo and possibly Sterkfontein. Body size is poorly known and degree of dimorphism is unclear but some individuals weighed 40-50 kg.
Brain size ranged between 650-800 cc. and an endocast shows some frontal lobe features not seen in australopithecines and supposedly characteristics of Homo sapiens. It has been suggested that these imply speech capabilities. The brain case is rounded, and resembles an enlarged, allometrically scaled version of A. africanus.
The face was large and deep, with big inceasers and canine and large cheek teeth . wear patterns on molar and pre molar broadly resemble those of Australopithecus. Post cranial are difficult to assign, but an innominate, femur, and tibia are probably habilis. The pelvic and leg bone differ from homologues of Australopithecus and resembles those of H. erectus, being similar to modern human bones, but more robust.
They imply a positional repertoire, gait, and ranging pattern more similar to humans than to australopithecines.
General Characteristics
- The cranial capacity ranges between 650-800 cc. The cranial capacity was greater than that of australopithecines and approached that of H. erectus. The brain to body size ration also suggested evolutionary advancement.
- Unlike australopithecines the frontal lobes (seats of mental ability) of brain were well developed they almost approximated that of modern man. The cranial vault was rounded.
- Hominid dental structure : the premolars are particularly humanlike.
- The mandibles (lower jaw) are comparatively less massive than that of australopithecines. However the dental arcade was not parabolic.3
- The Hind limb morphology clearly approached the human foot.
- H. habilis was a tool maker. The tool kit is referred to as "Olduwan Industry." It included cutting tools, scraping tools and tools to make tools. Meat, plant and wood were worked upon. Bones, seeds and nuts were cracked open. There is an evidence of camp sites also.
- Larger brain involves gestation, larger new born and greater longevity. The brain enlargement was perhaps the cause and the effect of dietary shifts in H. habilis.
The distribution of H. habilis and P. boisei overlaps. Both are found in east Africa during 2-1 million year ago. Homo habilis originated from some gracile australopithecines and represents a massive increase in brain size in short span of time. Some scholars disagree to the view that these are true Homos. Their teeth are large relative to their body size and their limb proportion shows that they are closer to australopithecines than Homo. Some even suggests placing these fossils in separate genus.
PHYLOGENETIC IMPLICATIONS
There has always been a question haunting the student of paleo-anthropology, why homo habilis, not aethiopithecus habilis?
One of the main criteria of for inclusion in the genus Homo was brain size. Different authorities had different threshold sizes, ranging from 700-800 cc. Most australopithecines were around 500 cc, while Homo erectus was above 1000 cc. What of H. habilis?
For some authors the brain size was more than 600 cc would suffice for inclusion in homo, along with the other feature indicated by the fossils, such as upright posture and bipedal walking and the precision grip in the hand. The first Homo habilis specimen had a brain size estimated at 680 cc.
With the discovery of Olduwan tool tradition, the discussion centered on tool making. Many authorities were unhappy about tools as admissible evidence in support of this new species. The history of hominid fossil discovery is littered with names coined in excitement of a new find. Many of these names have not stood the test of time.
In the mid-1960s many people thought it probable that homo erectus had evolved from some type of australopithecine. Therefore it would be natural to find fossils representing transitional forms-the result of evolution in action. Rather than create a separate species, this new form should be placed either with its predecessors, the australopithecines or with its successors, Homo erectus. According to Le Gros Clark, Homo habilis can be easily accommodated within A. africanus.
There were plenty of other opinions about where Homo habilis fitted into the hominid evolutionary tree. Leaky long held the view that our modern species, Homo sapiens was in fact very ancient and could be direct descendent of homo habilis. This would push Homo erectus out on to a limb, making it our cousin and not our ancestor.
In 1972 Richard discovered a skull at Koobi For a which had a cranial capacity of 800 cc . A brain size of this surely meant the creature to be member of genus Homo ?
Till today more than 20 Homo habilis individuals have been found. Besides having a bigger brain than australopithecines, Homo habilis also had a higher brain to body ratio compared to apes, the temporal lobes which deal with memory and other mental functions are more developed; so are the parietal lobes which analyse information coming in from senses and he speech processing area is also developed.
The version of our evolutionary tree in which A. afarensis gave rise to H. habilis which evolved into Homo erectus, which in turn Homo sapiens has an appealingly neat straight line feel to it. But standing back from the main habilis time period about1.9 - 1.6 million years ago brings various complications.
What about before the habilines? An alternative to the afarensis-habilis-erectus-sapiens view is that "early Homo" was already living more than 3 million years ago. Gradually the early homo became Homo sapiens. Whether it passed through the habilis stage on the way is an area lively debate.
What about after the habilines ? by 1.6 million years ago, another human species had appeared the taller, long legged, bigger brained Homo erectus. If habilis did evolved into erectus, could such a great evolutionary change have taken place so quickly ? one possibility is punctuation- a rapid burst of change in the type of evolutionary process known as punctuated equilibrium. Another is that some relatively isolated habilian populations began to evolved into erectus at much earlier date, leaving the rest of their kind to continue unchanged, and eventually go extinct about 1.6 million year ago.
The time between 2 and 1.5 million year ago has been called the crucial humanizing period. Recent fossils find from this time have widened the debate as to how many species of hominids lived in Africa then, and who evolved into what. More fossils may serve to clarify the picture ot further to confuse it.
Homo erectus
The study of fossil primates is the most essential requirement in order to study the emergence and development of man. At the beginning of 19th century, with the emergence of paleontology, comparative anatomy geology etc. the relations of man's antiquity were established by studying the fossil evidence.
Fossils are relics or traces of a former living things or plants preserved in rocks that are unchanged in structure. In tracing the ancestry of man, physical anthropologists have concentrated their attention to fossil primates.
The quaternary period, especially the Pleistocene epoch may be regarded as the age of fossil remains viz. ape like man, early man and Homo sapiens.
Three major stages which have been recognized in relation with hominid evolution are as follows:
- Australopithecines stage.
- Homo erectus stage.
- Neanderthalensis stage.
Australopithecine fossils lasted from about 5.5 - 1.5 million years B.P. (before present). They shared bipedal locomotion with the Hominids. These fossils belonged to various parts of Africa. In South Africa, Australopithecus africanus group were discovered at a place called Taung and Paranthropus robustus at Kromdraai.
In East Africa also Zinjanthropus boisei and Homo habilis were discovered at Olduvai Gorge. Australopithecus was again discovered from West Africa. Among these fossils, it is suggested that Homo habilis should be placed with the members of the later hominids.
Homo erectus:
The period between 2 to 1.5 million years B.P. (before present) provides reasonable points of transition from some of the Australopithecines to Homo. By about this time skull and jaws of Robust australopithecines were found indicating that this robust type continues to exist after the evolutionary appearance of Homo erectus. And the period between 1.5 to 125,000 years BP is regarded as the transition from Homo erectus to Homo sapiens. Under Homo erectus, different generic names such as Pithecanthropus erectus, Sinanthropus pekiinensis and Atlanthropus mauritanicus are placed.
The specific name Homo erectus derived from Pithecanthropus erectus was given to the finds from Indonesia in the 19th century and when they were sufficiently distinct from human, they were given a separate generic name known as Homo erectus. The term Homo erectus means erect man.
Homo erectus fossils have been discovered in many parts of the world, including Java, China and North, East and perhaps South Africa but rarely from Europe. In North Africa, three complete lower jaws and a piece of skull cap similar to that of the fossils found in China were discovered.
In East Africa from Olduvai Gorge, Marry Leaky found a brain case that looks like Asian Homo erectus. In South Africa, a part of a skull and face were discovered at a site called Swartkrans. At this site, robust Australopithecines were accumulated at one time.
Earliest fossil finds are molar tooth, a skull cap (1891) and a femur (1892) of Homo erectus were described by Eugine Dubois. The finds were discovered on Sole river in central java. Before this discovery Ernst Hackel, a German scientist had proposed that if and when a link between apes and man is found it should be named pithecanthropus (ape man) the finds were accordingly named Pithecanthropus erectus.
Another juvenile skull (of a child, less than 2 years in age) that bore striking resemblance with the earliest found fossils, and discovered by H. R. Von Koenigswald in eastern Java was given the name of Homo modjokertensis.
Soon after, two skull caps of the species were discovered at a near by site, Koenigswald believed that the fossils closely resembled those of the Pithecanthropus erectus. During world war II Japanese took over Java, Koenigswald was made a prisoner and fossils were confiscated. Meanwhile, in anticipation of all this he had already distributed the casts of the fossils to his friends. Fortunately, to his pleasant surprise Koenigswald was released after the war and all his belongings including the wealth of fossils were returned to him.
But the fossil finds of china met different fate. The first fossil of the species, a few teeth, found at Zhou-kau-dein between 1921 to 1927 after thorough study by D. Black were assigned the namw of Sinanthropus pekintensis. Black had realized well that the species was different from our own. In 1929 W.C. Pei, for the first time, discovered the skull of Sinanthropus.
By 1937 the site had yielded skulls, teeth and other fossil bones of about 45 hominid individuals, along with stone tools, bones of thousands of animals and evidence of fire.
The area in 1941 was under Japanese control while the fossil expedition was being undertaken by Americans. Sensing a threat to survival of fossils, the fossils were packed and shipped to U.S.
But as ill luck would have it the sip was captured, the fossils disappeared never to be seen again. Today whatever we know of Zhou-kau-dein fossils is based their casts prepared before this mis-happening. The Homo erectus name was given by Beuttner Janusch as late as 1960s to all fossils mentioned above. Till date H. erectus fossils have been found in Indonesia, China, Algeria, Morocco, Tanzania, Germany and Hungry.
European fossils are however awaiting authenticity. The earliest fossils describes by Dubois is popularly known as Java-man. The man was an individual who stood tall at 5.8 inches. The skull cap is primitive with heavy brow-ridges and with a cranial capacity of 750-900 cc (between ape and man). Femur bone a well-developed linea aspera.
Most information of Homo erectus we have today is however based on14 skulls 147 teeth and a few post cranial fragment of about 45 individuals made available from Zhou-kau-dein caves which H. erectus inhabited for a thousand years or so. The fossil finds of Java popularly known as java man have been given the name of H. erectus.
While those of the China have been placed in a separate sub species of H. erectus pekiinensis. The two sub species of erectus differ only marginally. The skull vault of pekiinensis. Is slightly higher and brow ridges tend to curve more. In lateral view pekiinensis forehead rises at a steeper angle than in its erectus counterpart.
Compared to Peking Man, Java Man has more massive jaws and teeth. Following information belongs to H. erectus as a species and not to any individual fossil.
ANATOMICAL FEATURES:
- Optic lobes of brain are less developed than man,
- Parietal lobes comparatively well developed (power of articulation of Thoughts);
- Cranial capacity between 925 cc (in 8 or 9 year old child) ad 1225 cc (within human range);
- Jaws and teeth large with space between teeth,
- Dental arcade more parabolic than apes,
- Science of taurodontism (enlarged pulp cavities; also found in Neanderthal and some modern Man) upper inceasers showed shaped with sides seeming to curve inwards (mongolide traits),
- No diastema or chin, nose broad and low ridges;
- Man above five feet and women slightly less tall.
- In post cranial anatomy erectus resembled modern man.
Cultural Advancements:
Homo erectus was the first perennial tool makers. Tool bearing sites spread from north-west Europe to south east Asia are much more numerous than the sites yielding fossils. The stone tool culture of mid-Pleistocene origin have been generally ascribed to Chellean and Acheulian traditions. Choppers and chopping tools found mainly in China are restricted in distribution up to central Asia, while hand axe is the most common tool in other parts of the world.
H. erectus, in the absence of any defence organ used these tools for hunting. Hand axe was a multipurpose tool that could be used for cutting, chopping, scrapping, boring etc. and throwing at the animals. So essential was the use of stone tools that at Zhou-Kou-Dien stones for making tool were brought from a distance of more than 3 kilometers.
The earliest evidence of news of fire by erectus is seen in China some five lakh years ago and much later at Vertess-zollos in hungry Terra Amata in France. Fire could be used in a number of ways. It provide security, protection from predators, warmth at night and of course light. It provided for a group center and increase the working capacity in terms of working hours.
It also helped in splitting stones from bigger rocks . only fire could sustain erectus in colder regions like in northern china. In caves sites where fire has been located presence of charred bones indicates that (at least occasionally) fire was used to cook food. Homo erectus was a hunter gatherer. At Torralba in Spain, Howell has discovered a mud hole that was used to trap big animals.
Here the bony remains of more than 40 elephants bear testimony to the hunting skills of middle Pleistocene hunters. Similarly at Olorgesalie in Kenya remains of more than fifty giant baboons (as a troop) are found that must have been cornered by homo erectus. Nowhere has been found the evidence of deliberate burials and we feel handicapped in understanding the rituals, if any, of this early homo species.
How did erectus communicate? Had he developed language?
These questions cannot be answered in yes or no. We do not have the brain of Man's ancestor available with us. The facts are however inferred through the study of available skulls, crania and skull caps. Traditionally man's large brain has been given the credit of development of speech.
It is not true otherwise the whale and elephant too could have enjoyed this fantastic gift of nature. Even in brain body weight ratio, capuchin monkey possess one gram of brain for 17.5 grams of body the same ratio for man is 1:44. The brain body weight ratio too is not a good indicator in this regard. The development of brain therefore should be understood in terms of its use and not as a proportion to body weight or its overall size.
Major areas in brain are present in man, apes and some other mammals alike but none other than man can communicate through a verbal language. Animals communicate through symbols and gestures. Australopithecus with the development of brain in the range of that of great apes does not seem to have had a language.
It seems that development of brain for this purpose has been the result of group hunting and social life over millions of years. This could easily have resulted in the development of memory areas in the occipital lobe as also the areas of ideas in the frontal lobe. The development of language in some people followed by the selective advantage conferred on them must have resulted in the spread of lingual communication.
The development of different motor areas through a regular practice of stone throwing and game play too must have contributed in their own way. The development of language is followed the progress in tool technology. The modern complex language is however attributed to the arrival of modern man.
HOMO ERECTUS-PHYLOGENETIC POSITION
Though H. erectus has been placed in genus Homo along with modern man, he exhibited many primitive characters as given below:
- Bones of cranial vault - very thick
- Strongly developed supra orbital torus extending above, the orbits as an un interrupted bar of bone
- Receding frontal bone
- Well-developed occipital ridge that extended into the supramastoid region and produced an angular contour to the occipital bone.
- Small mastoid process
- Greatest width of cranial at the level of ear lobes
- Broad nasal bones
- Pronounced alveolar prognathism
- Massive body of mandible
- Frequent occurrence of multiple mental foramena.
- Larger upper inceasers.
He however exhibited several characters which place him closer to modern man compared to his predecessors these are:
- Cranial capacity overlaps the lower range of that in H. sapiens and the cranial vault is inflated.
- Increased flexion of face on the brain case (compared to Australopithecus) so that the anterior cranial fossa extends well over the orbits.
- Relative size of face reduced more than in Australopithecus
- Foramen magnum positioned more anteriorly than in Australopithecus
- Conformation of tempero-mandibular joint as in H. sapiens
- Dental arcade parabolic in shape
- Dental morphology more like H. Sapiens than Australopithecus
- Limb bones indistinguishable from those of H. sapiens in shape and size.
HEIDELBERGENSIS
On October 21, 1907 was discovered a mandible that is now popularly known as Mauer jaw or Homo heidelbergensis. The discovery was reported by Otto Schoetensack, a professor of anatomy in Heidelberg University in 1908. The fossil was discovered was 65 feet below ground level. It was the first human fossil material recovered from middle Pleistocene of Europe.
Ascending rami being extremely broad (60mm against an average of 37 mm in modern man) appear square. Upper surface of ramus (mandibular notch) is comparatively shallow. Coronoid process is present at a lower level than condyle and is blunt (similarity with gibbon).
Horizontal rami are large with a maximum thickness (23 mm against 14 mm in modern man) at level of third molar the symphysis has a convex and receding curve externally (like apes unlike man) and is quite thick (17 mm).
Fifth cusp, though small, is present in all the molars and there are large pulp cavities. With this combination of simian and human features, Schoetensack concluded that the fossil represented a stage between ape and man. Duckworth suggested the inclusion of Mauer jaw in genus pithecanthropus. While opposing Duckworth views Boule and Valloi consider it as an ancestor of Homo Neanderthalensis of western Europe.
Presently, however the jaw is placed by some in homo erectus with a different sub species of heidelbergensis; while some others consider it as early Sapiens.
The Mauer deposits have been dated to about three lakh sixty thousand years ago and this reflect the age of Mauer Jaw.
Transtion to Homo Sapiens
Genus Homo evolved about 2 million years ago and erectus was probably the first Homo species. It however took erectus over one and half million years to evolve into the modern species of sapiens. The cave of Arago in Pyrenees Mountains of southern France have yielded a skull and several mandibles belonging to the ending phase of the middle Pleistocene, Riss glaciation, which belong to individuals more advanced than homo erectus but less so than the sapiens. In Java where the fossils of erectus and sapiens have been unearthed, recent discovery of some skulls at Sangiran clearly speaks of the continuity of erectus into sapiens. The fossils have been placed intermediate between the two species.
The Swanscombe skull found near river Thames, England is less than half a million years old. The skull walls were thin than Beijing Man, the inion is is below the opisthocranion resulting in rounder contour of the skull. The maximum breadth of the skull lies not near the base but higher up on the parietals. The pattern of mid-mengeal vessels, is more complex, all these features put Swanscombe skull more close to homo sapiens.
Another middle Pleistocene skull similar to the Swanscombe skull has been discovered in Steinhein, near Stuttgart, Germany. Popularly known as Steinhein Skull, it has walls further thin in the range of homo sapiens while the cranial capacity approx. 1185 cc is less than that of Swanscombe skull which is approx. 1300 cc.
Other features in the two skulls are similar. The two skulls discovered at Fontechevade have features more advanced than the Swanscombe or the Sreinhein skull. These fossils belonging to upper Pleistocene with a cranial capacity of about 1450 cc and absence of thick brow ridges are further closer to the sapiens. All the above mentioned fossils belonged to the transitional phase of the Human evolution. The fossils are marked by greater cranial capacity, greater development of skull intermediate between H. sapiens and H. erectus, and nuchal lines below the opisthocranion. The fossils represent represents a form ancestral to homo sapiens. Obliviously, H. erectus during the later mid-Pleistocene and early upper Pleistocene was evolving into H. sapiens.
NEANDERTHAL MAN (HOMO NEADERTHALNSIS)
Neanderthal Man derives his name from Neanderthal (Neander Valley) Dusseldorf, Germany, where his fossils were found and described first.
The earliest fossils of Neanderthal Man comprising of a skull cap with upper margin of the orbits, parts of pelvis, two femurs, two humeri, two Ulnas, five ribs and one each of radius clavicle and scapula were found in Neanderthal, Dusseldorf, Germany. The individual, an adult had an estimated cranial capacity of 1230 cc . Post cranial parts so heavy musculature marks but structurally are exactly like the present - day bones. Today Neanderthal fossils are known from Spain, France, Italy, Germany, Slovakia, Hungry, England, Yugoslavia, Israel, Iraq, Libya, Lebanon, Morocco , Russia, Java, China and Pakistan.
His cultural finds however are much more numerous and have been found in many other countries of the world. Most of the fossils finds belong to the last interglacial and last glacial. The individual is worthy to be placed in the evolutionary ladder between Homo erectus & Homo Sapiens.
Anatomical Features
The main features of Neanderthal anatomy are:
- Increased brain size & large capacity.
- Dolichocephalic, skull wall and brow ridges thick
- Heavy chinless Jaw.
- Some facial prognathism forwardly projected upper jaw.
- Frontal part not fully developed.
- Comparatively broad nose
- No canine fossa (Unlike man) instead slight puffing out of the bone region.
- Heavily muscled thick neck.
- Barrel chested with powerful muscles of legs and arms
- Sylvan fissure (expression of language) in brain well developed but less developed pharynx.
- Stature 5 feet or more.
Cultural Advancement:
- The first known species to have control of fire about 50000 years ago.
- The choppers and chopping tools were replaced by the use of hand axe in Europe and Africa.
- In Asia however OVATE cleavers become popular.
- In Africa there is continues evolution of tools from crude, heavy Chellean hand axe to better made Mousterian hand-axes.
- Flake tools become more and more common with time, most common techniques for getting flakes being prepared/core and Levalloisian.
- The most common flake tool was the "Point" probably used by hafting into the end of a spear.
- Scrapers were also commonly used. The blades, found in low frequency in the early stage become more common in the later period & probably continued into upper Palaeolithic of modern man.
- Probably had strong social organisations, otherwise La Chappelle man as also the handicapped one of the Shanidar could not have survived in old age.
- With Neanderthal we find the first direct evidence of prevalence of rituals as also the deliberate burials.
- In Tashik-Tash (erstwhile USSR) the burial of young Neanderthal was surrounded by wild goat skulls with horn pointed towards the burial.
- In Monte Cicero (Italy) the skull has been broken from the base to extract brain.
- Tools are most commonly found associated with the dead. At Shanidar too, the association of dead with arrangement of bear skulls as also the floral offerings indicates some form of primitive religious practices.


